

Response to Existing Biochemical Autism Pathology Models
Response to Existing Biochemical Autism Pathology Models
The distinction is not whether biochemical changes exist in autism. The distinction is how those changes are organized into a system-level cascade and what governs their behavior over time.
Recognize biochemical involvement
Many models examine metabolic, immune, mitochondrial, redox, and neurotransmitter pathways independently or in clusters. Each existing model has a unique focus.
Defines the organizing mechanism
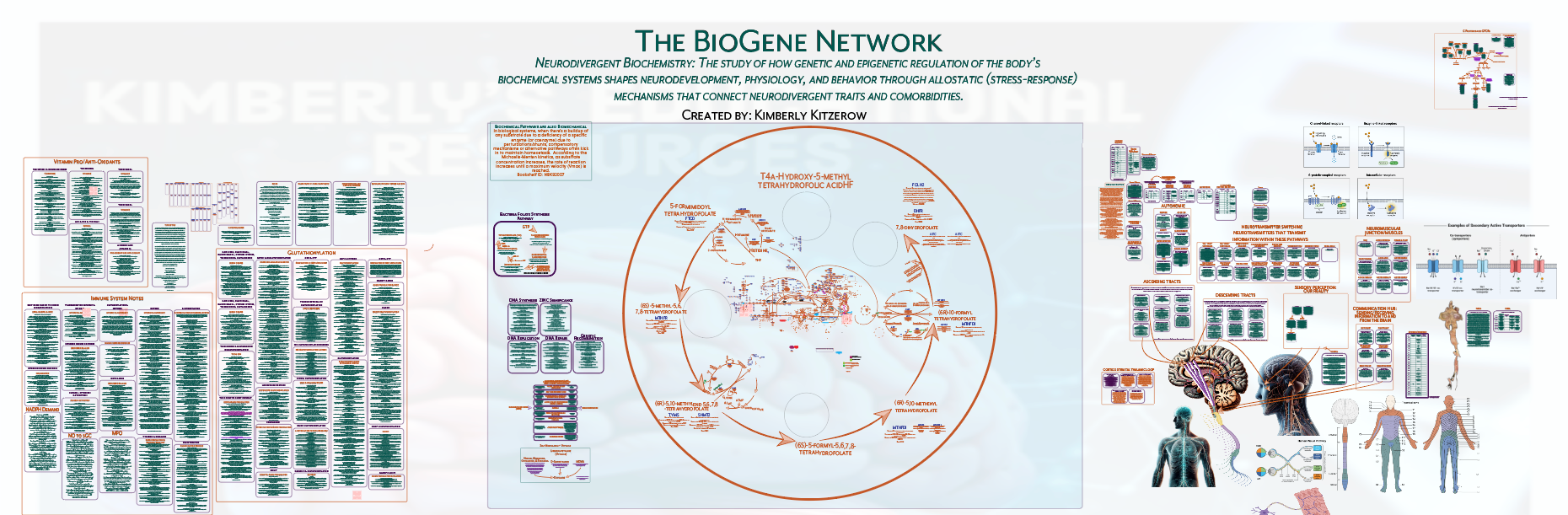
BH4 shunting is positioned as the central organizing mechanism that structures how these biochemical changes converge into a unified cascade.
What Determines Cascade Behavior
The cascade is not defined by mechanism alone. It is shaped by how regulatory systems are activated, how long they remain active, and how that activation interacts with temporal system domains.
Type of Activation
Situational, chronic, or genetically sustained activation.
Duration
Short-term versus prolonged activation determines system load and downstream impact.
Regulatory Systems
Immune, metabolic, nervous system, cellular repair, and genomic regulation.
Temporal Domains
Ultradian, circadian, developmental, and lifespan timing influence how changes accumulate.
Kitzerow's contribution is a unified cascade model in which BH4 redistribution operates as the organizing mechanism and system behavior is determined by duration, type of activation, and timing across regulatory domains.
How Kitzerow’s Autism and the Comorbidities Theoretical Model Relates to Existing Biochemical Autism Models
These models do not operate from the same organizing logic. Kitzerow’s Autism and the Comorbidities Theoretical Model differs not by recognizing biochemistry, but by how the cascade is structured, centralized, and explained.
Amy Yasko
Organizing Logic
Organized around inherited polymorphisms affecting methylation, transsulfuration, detoxification, and related pathway checkpoints.
Primary Focus
How SNPs, toxins, and pathway inefficiencies alter biochemical throughput and create intervention points across interconnected pathways.
How Kitzerow’s Framework Differs
Kitzerow’s Autism and the Comorbidities Theoretical Model is not organized around pathway inefficiencies or SNP-driven checkpoints. It is organized around BH4 shunting as a constrained regulatory state, with methylation and transsulfuration changes interpreted as downstream effects within a coordinated cascade shaped by system activation and duration.
Richard Frye
Organizing Logic
Organized around biomarker dysregulation, metabolic findings, and therapeutic response across mitochondrial, folate, methylation, redox, and related domains.
Primary Focus
How measurable biochemical abnormalities and treatment-responsive patterns can inform autism-related metabolic and mitochondrial dysfunction.
How Kitzerow’s Framework Differs
Kitzerow’s Autism and the Comorbidities Theoretical Model does not organize findings as parallel biomarker abnormalities. It organizes them as outputs of a single cascade, where mitochondrial, immune, redox, and methylation changes emerge from BH4 constraint within a unified, duration-dependent system.
Sharon Meglathery
Organizing Logic
Organized around the RCCX module and the physical proximity of linked genes on chromosome 6.
Primary Focus
How inherited variation in closely linked genes may jointly influence neurodivergence and overlapping stress, immune, endocrine, and connective tissue patterns.
How Kitzerow’s Framework Differs
Kitzerow’s Autism and the Comorbidities Theoretical Model is not organized by gene proximity. It is organized by biological function, where genetic variation is interpreted through its effect on regulatory systems that converge on BH4 redistribution and cascade behavior over time.
Kitzerow’s Autism and the Comorbidities Theoretical Model
Organizing Logic
Organized around BH4 shunting as the governing mechanism structuring a unified biochemical cascade.
Primary Focus
How regulatory system activation, temporal domains, duration, and source of activation shape cascade behavior and the emergence of traits and comorbidities together.
What Makes It Distinct
Kitzerow’s Autism and the Comorbidities Theoretical Model is organized as a system-level cascade. BH4 shunting serves as the organizing mechanism, while regulatory system activation, temporal domains, duration, and source of activation determine how traits and comorbidities emerge together.